- Drift in biology is “evolution by randomness”. The ‘drift’ idea was championed by Motoo Kimura as “The Neutral Theory of Molecular Evolution”. Kimura was motivated by JBS Haldane’s dilemma: too much polymorphism exists for “natural selection” to be able to pick and chose “beneficial” genes due to gene colocation in the genome and to the propagation time in the population. Of course, Haldane’s dilemma is only due to the conflict between his nonnegotiable assumptions that “natural selection”, “fitness” and “beneficial mutations” are all real and the observable realities of biology. The ‘drift’ idea claims that random events cause allele frequencies to change over time and in extreme cases, ‘drift’ is seen eliminating or fixing certain alleles in the whole population. Drift is supposed to operate independent of “natural selection” to the extent the allele is not strongly “beneficial” or “detrimental” for its frequency to be primarily a product of “natural selection”.
- Outside of “evolution”, drift adds to nothing or is aimlessly carried by known forces. In the first case it has no effect on anything either in the short or the long term. In the second case it is explained by the carrying force and therefore is itself not an explanation of anything. Examples of driven drifts are driftwood, wind-driven snow, rain, cloud, dust, or smoke as well as aircraft moved by air currents (all these driven). Inconsequential drift examples include aimless course or random movement. In biology too, genetic drift should either be inconsequential or, if lasting changes are observed, not drift but the real driving forces behind this drift should be held responsible.
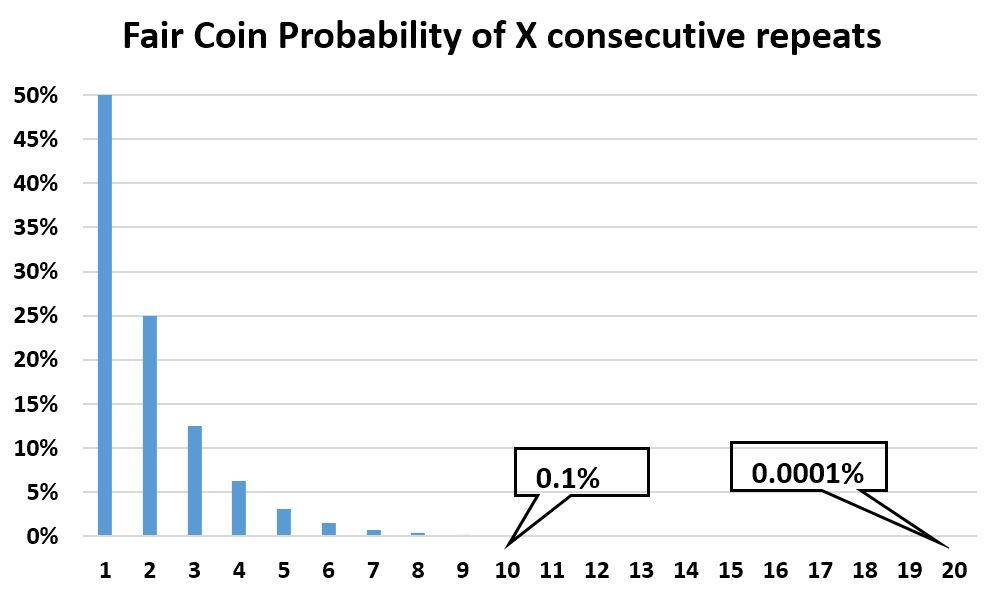
- A fair coin is a good model for inconsequential biologic drift. But how can that be, given mutations can take the population in a myriad directions, not just two (head and tails)? The fair coin model works because from a starting population, there are only two mutually exclusive directions possible on any of the myriad dimensions: towards or away from the initial value. For instance, mutations can make a second generation taller, greener, heavier, etc. But the third generation can only be even more tall, green, heavy, etc. than the original population, or less so than the second generation and therefore more like the original population. Just as with a fair coin, the output is binary: further away (call it ‘tails’) or closer to (call it ‘heads’). To prove random drift irrelevant in biology, let us consider the statistical case for drift by using a fair coin sequence. Yes, there is a statistical probability however small to see a series however large of tails (or heads), but eventually that series must end. As the chart shows, the probability of a sustained random drift decreases exponentially to nil. And over a much longer time, the realized events still sum up to an average of “no change”. This is easily verifiable experimentally. And if a significant bias persists over the long term, then the coin cannot possibly be ‘fair’.
- Population bottleneck cannot lead to “evolution”. Populations sometimes decrease dramatically due to competition, disease, predation or natural catastrophes. But killing the majority of one allele while sparing most of another is either a non random scenario or not possible except in the tiniest of populations and thus inconsequential. Post bottleneck, mutations occur and are claimed to further cause the strain to diverge from the original. But if no driving force is involved, there is no reason to believe mutations would work in a diverging direction only as the statistics discussion proves. Therefore, only a driving force, not drift can be responsible for long term changes following a bottleneck event. Alleles could indeed disappear from a population due to a series of random events, but either the population is small hence the loss is irrelevant; or, if that small population grows, a mutation will likely reintroduce the allele; or, in an already large population, the allele loss is highly unlikely due to random events not scaling up with the population size. Either way, random allele loss cannot possibly drive “evolution”.
- Founder effect also cannot lead to “evolution”. Many islands contain unique species due to having been populated by only a few organisms. Similar to bottlenecks, founder effect reduces the amount of genetic diversity in a population, meaning certain biological characteristics are overrepresented and these individuals are hypothesized to form new populations with different gene pools from the original population. But the same problem arises: the sum of mutations cannot be divergent unless a force other than drift itself causes that. If the environment weren’t drastically different, rather than diverge one would expect the new population to converge to the old strain instead given that at least that strain is known to be viable. Furthermore, if one small group found its way to the island, what prevents other small groups from repeating the trip and cancelling any such founder effect? The best explanation for the founder effect is thus that it is an ephemeral artifact and that all lasting differences are just adaptations unrelated to any random drift.
- There is more to the “dilemma” than Haldane’s take, and it’s all bad for “evolution”. A few things we know for sure: a) bottleneck populations are much more homogeneous as observed in the northern elephant seal, European bison, domestic animals, etc.; b) unique founder features (including injurious ones) persist for a long time in the founded population c) established populations (no bottleneck) are likely to be polymorphic with morphs in long term equilibrium. The theory of “evolution” predicts “beneficial” mutations taking over the population resulting in an “evolved”, better population over time. This is inconsistent with polymorphism and with the propagation of the founder’s features (some seeming “deleterious”) over time. Not only that, but normal population variability argues against “natural selection”. Because “fitness” functions cannot be calculated even in principle, let alone forecasted for any of the morphs, a hypothesized “natural selection” cannot possibly operate in biology. Furthermore, newfound populations, while adapted to their new environment, are in no way superior to their mainland cousins, even after the longest time from the split. This clearly shows “beneficial mutations” impossible. Thus, Haldane’s dilemma can easily be solved by removing his prejudiced, hopeless and disproved assumptions of “natural selection”, “fitness” and “beneficial mutations”.
- Balancing selection, an alternative to drift, fails to solve Haldane’s dilemma. The big idea of “balancing selection” is that the “fitness” of an organism with two alleles (heterozygote) is higher than the “fitness” of either allele (homozygote). Perhaps “fitness” changes in time from young to adult. Or “fitness” of a phenotype is dependent on its frequency relative to other phenotypes – rare morphs of prey are actually “fitter” due to predators concentrating on the more frequent morphs. However, there is no way to forecast anything of this sort or even to explain with any confidence the frequencies of these alleles. An elusive, fluid “fitness” attempting to explain all ends up explaining nothing, therefore it is a redundant hypothesis. And why would “fitness” changes in time? Why wouldn’t a morph “evolve” to do away with this temporal weakness? And what should the frequency and number of alleles be? Again, no rhyme or reason. Take ABO blood system. Why not more or fewer types? Why these frequencies? And why the current distribution? These are not just unanswered; they are unanswerable question with any degree of confidence.
- In conclusion, both drift and “natural selection” are disproven by polymorphism. As seen, drift in itself is not an explanation. All bottleneck drift “examples” are of theoretical cartoonish type (colored balls/ rabbits/ bugs). Some founder effect examples are real but inconsequential (northern elephant seal and Amish polydactyly) as clearly none of these leads to any “evolution”. Meanwhile, “natural selection” is perplexed and for sure inexistent as “fitness” is nowhere observable let alone quantifiable, therefore also a chimera.
Pro-Con Notes
Con:
Pro:
Links:
https://en.wikipedia.org/wiki/Haldane%27s_dilemma
https://en.wikipedia.org/wiki/Neutral_theory_of_molecular_evolution
https://en.wikipedia.org/wiki/Genetic_divergence
https://en.wikipedia.org/wiki/Balancing_selection
https://en.wikipedia.org/wiki/Heterozygote_advantage
https://en.wikipedia.org/wiki/Polymorphism_(biology)
https://en.wikipedia.org/wiki/List_of_polymorphisms#Ants
https://en.wikipedia.org/wiki/Heterozygote_advantage#Cystic_fibrosis
https://sciencing.com/difference-between-natural-selection-descent-modification-17942.html